|
|
|||
|
|
|||
|
|
|||
|
Il termine sensibilità propriocettiva venne introdotto nel 1906 da Sir Sherrington per indicare l’insieme dei segnali nervosi che si originano durante il movimento. Secondo questa definizione, gli stimoli adeguati sono applicati dall’organismo stesso, e di qui il termine “proprio”, a recettori specifici che si trovano all’interno degli organi di movimento. A distanza di circa un secolo, tale definizione appare ancora attuale, anche se ha subito delle estensioni. In origine si annoveravano nell’ambito della sensibilità propriocettiva soltanto i segnali provenienti dai fusi neuromuscolari e dagli organi muscolotendinei del Golgi. Attualmente vengono inclusi nella s.p. anche stimoli provenienti da altri recettori, tattili, pressori, dolorifici, ecc. I principali e meglio studiati tra i propriocettori sono i fusi neuromuscolari e gli organi muscolotendinei del Golgi. Fanno parte dei recettori da stiramento, perché sensibili alle modificazioni fisiche indotte nel muscolo dallo stiramento delle fibre muscolari stesse. |
|||
|
|
|||
|
Struttura e funzione dei fusi neuromuscolari |
|||
|
|
|||
|
|||
|
Le fibre muscolari modificate che compongono il fuso vengono definite intrafusali e sono più sottili delle fibre muscolari vere e proprie. Di solito si distinguono due tipi di fibre intrafusali : |
|||
|
|||
|
E’ possibile, inoltre, riconoscere nelle fibre a sacchetto di nuclei due sottotipi :
|
|||
|
Esse sono notevolmente diverse tra loro sia come composizione ultrastrutturale, che come caratteristiche biochimiche, sia per le loro proprietà meccaniche. L’innervazione del fuso è poi assai complessa e consiste di fibre sensitive e motorie. Le fibre sensitive afferenti si distinguono in due gruppi :
|
|||
|
Le fibre afferenti primarie, ad alta velocità di conduzione, con diametro tra 12 e 20 micromm, appartenenti al gruppo Ia della classificazione delle fibre nervose di Lloyd, originano da terminazioni a spirale che si avvolgono sulla porzione centrale delle fibre intrafusali di tutti e tre i tipi descritti. Le fibre afferenti secondarie, di minore velocità di conduzione, e di più piccolo diametro, appartenenti al gruppo II della classificazione delle fibre nervose di Lloyd, provengono prevalentemente da terminazioni a spirale avvolte sulla porzione paracentrale delle fibre a catena di nuclei. A queste si aggiunge il contributo di terminazioni a fiorami, distribuite quasi esclusivamente sulle fibre intrafusali a sacchetto di nuclei. Una caratteristica particolare del recettore fusale, consiste nella presenza di fibre motorie provenienti dai motoneuroni gamma delle corna anteriori del midollo spinale, che si distinguono in due tipi: |
|||
|
1. fibre gamma dinamiche, che, in numero di 1 o 2 per lato, innervano le porzioni polari delle fibre “a sacchetto di nuclei dinamiche; 2. fibre gamma statiche, che innervano sia le fibre intrafusali “a sacchetto di nuclei statiche”, sia le fibre a catena di nuclei. |
|||
|
|
|||
|
Per comprendere il meccanismo con cui il fuso riesce a segnalare le variazioni di lunghezza del muscolo, bisogna ricordare la disposizione anatomica ed istologica dei fusi nell’ambito del tessuto muscolare. La capsula del fuso si fonde, ai suoi due poli, con l’endomisio, la guaina connettivale che riveste le fibre muscolari extrafusali. Questa disposizione in parallelo, permette che ogni allungamento del muscolo si ripercuota anche sulle fibre intrafusali, così come lo stiramento produce un distanziamento delle spire della terminazione nervosa avvolta sulla porzione centrale delle fibre intrafusali, là dove si trova il fluido gelatinoso che, molto probabilmente svolge un’azione lubrificante e permette degli spostamenti indipendenti tra loro delle varie fibre. Dobbiamo inoltre ricordare che i fusi neuromuscolari sono recettori di primo tipo. Recettori, cioè, capaci di rispondere alle variazioni di energia incidente con un potenziale elettrico chiamato Potenziale Generatore (PG). Lo stimolo produce una variazione elettrotonica del potenziale di membrana molto simile al potenziale post-sinaptico eccitatorio. Esso dipende direttamente dall’ampiezza dell’area stimolata e dall’intensità dello stimolo, ed è dovuto ad una modificazione non selettiva della permeabilità della membrana a tutti gli ioni. Sono soprattutto gli ioni Na+ ad entrare, spinti dal doppio gradiente, sia elettrico che chimico, creando un flusso intracellulare di cariche positive depolarizzanti. E’ proprio la deformazione che innesca il processo di trasduzione meccanoelettrica e la depolarizzazione della terminazione con l’invio del potenziale d’azione al 1° nodo di Ranvier della fibra afferente. La sensibilità del fuso allo stimolo di stiramento è molto elevata, bastano piccolissime variazioni di allungamento del muscolo per far variare significativamente la frequenza di scarica delle fibre afferenti. Grazie alla presenza dei due tipi diversi di fibre intrafusali, il sistema nervoso riceve informazioni sia sull’entità dell’allungamento muscolare attraverso le variazioni di scarica delle fibre secondarie, sia sulla velocità dell’allungamento attraverso le fibre di gruppo Ia. Data la disposizione in parallelo dei fusi, essi subiscono, durante la contrazione delle fibre extrafusali, un accorciamento che inibisce e fa cessare del tutto la scarica afferente. Il controllo sulla sensibilità del recettore è operato dal sistema di innervazione gamma ed alfa. L’innervazione gamma produce una contrazione localizzata alle estremità polari delle fibre intrafusali, che, a sua volta, determina uno stiramento della loro parte centrale, dove sono avvolte le terminazioni nervose. In questo modo la detenzione del fuso prodotta dalla contrazione delle fibre extrafusali ad opera dei motoneuroni alfa, può essere compensata dagli effetti opposti prodotti dalla contemporanea attivazione dei motoneuroni gamma. In questo modo la sensibilità del recettore può rimanere costante per ogni livello di contrazione muscolare, inoltre il controllo di sensibilità del recettore, operato dal sistema gamma, è in grado di esaltare selettivamente la sensibilità statica o dinamica del fuso, attraverso i due tipi di fibre gamma che lo innervano. Gli impulsi provenienti dai fusi raggiungono il midollo spinale ed eccitano i motoneuroni alfa ed i motoneuroni dei muscoli agonisti con una trasmissione monosinaptica. E’ questa la branca afferente del riflesso miotatico, cioè l’allungamento attivo o passivo di un muscolo provoca, mediante l’attivazione dei propriocettori fusali, una contrazione riflessa che tende a riportare il muscolo alla sua lunghezza originaria. Il riflesso miotatico è presente sia nei muscoli estensori che nei flessori, è particolarmente presente nei muscoli che si oppongono alla forza di gravità e svolgono un ruolo primario nel mantenimento della postura. In questi casi è la componente statica del fuso ad essere stimolata. Esistono centri sopraspinali di controllo dei gamma motoneuroni tonici, posti nella formazione reticolare e nel complesso dei nuclei vestibolari, che ottimizzano il significato funzionale del sistema propriocettivo dei fusi neuromuscolari. Diverso il ruolo delle afferenze secondarie, fibre di gruppo II, provenienti dalle terminazioni anulospirali poste sulle fibre intrafusali a catena di nuclei. Queste, accanto ad una funzione monosinaptica, esercitano complessi effetti polisinaptici di tipo eccitatorio sui muscoli flessori, di tipo inibitorio sui muscoli estensori. Le fibre di gruppo II, quindi, partecipano alla componente tonica del riflesso miotatico, in quanto sensibili agli allungamenti statici del muscolo; eccitano la componente flessoria dei motoneuroni spinali, attraverso connessioni polisinaptiche; in condizioni di ipertono, come la spasticità, la loro stimolazione inibitoria aumenta in funzione dello stiramento passivo esercitato sul muscolo. |
|||
|
|
|||
|
Struttura e funzione degli organi muscolotendinei di Golgi |
|||
|
|
|||
|
|||
|
di fibre muscolari, nell’uomo sono circa 20 le fibre muscolari di riferimento per ogni organo del Golgi. Le fibre muscolari poste in relazione agli organi muscolotendinei del Golgi appartengono a diverse unità motorie, ciò permette al recettore di campionare la tensione sviluppata da unità motorie di diversi tipi durante la contrazione muscolare. Le terminazioni nervose amieliniche di forma irregolare penetrano tra le trabecole connettivali del corpo tendineo. Lo sviluppo di tensione del muscolo stira i filamenti che diventano più rettilinei. Questa deformazione meccanica depolarizza i filamenti, innescando i potenziali d’azione nella porzione mielinica extracapsulare della fibra nervosa. Gli impulsi giungono al midollo spinale lungo fibre ad alta velocità di conduzione del gruppo Ib. Esse hanno un’azione inibitoria sui motoneuroni alfa e sui muscoli agonisti attraverso la mediazione di un solo interneurone spinale. Questi organi sono estremamente sensibili alla contrazione attiva dei muscoli e pressocchè indifferenti agli stiramenti passivi dei muscoli stessi. In passato si pensava che gli organi del Golgi fossero uno strumento di difesa dell’organismo per segnalare contrazioni eccessive e proteggere dalla rottura le inserzioni muscolari. Oggi, invece, si sa per certo che l’attivazione massiva di queste afferenze può arrestare la contrazione muscolare, ma la loro specifica funzione è quella di segnalare ai centri superiori la tensione media sviluppata da un selezionato gruppo di unità motorie, la frequenza di scarica aumenta con il progressivo aumento del reclutamento delle unità motorie. Questo meccanismo, oltre a rappresentare un meccanismo di controllo a feedback negativo per la contrazione, viene usato dall’organismo per la programmazione della forza di contrazione da parte dei centri motori. |
|||
|
|
|||
|
Proiezioni sopraspinali dei propriocettori muscolari |
|||
|
|
|||
|
I propriocettori muscolari inviano informazioni a varie strutture sopraspinali interessate alla programmazione ed al controllo sia della statica che del movimento. In particolare il cervelletto e la corteccia cerebrale sensitivo-motoria ricevono una cospicua mole di informazioni. |
|||
|
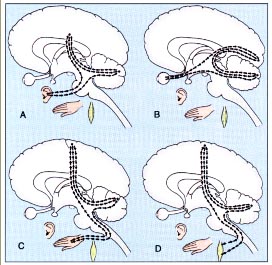
Le vie di afferenza dirette al cervelletto sono essenzialmente 4 : 1. tratto spinocerebellare dorsale, 2. tratto spinocerebellare ventrale, 3. tratto cuneocerebellare, 4. tratto rostrocerebellare. |
|||
|
|||
|
del Golgi sempre dagli arti inferiori. Le fibre del tratto spinocerebellare dorsale terminano sotto forma di fibre muschiose nel lobo anteriore del cervelletto. Il tratto spinocerebellare ventrale veicola nelle stesse regioni del lobo anteriore del cervelletto informazioni provenienti essenzialmente dai corpi muscolotendinei di Golgi e rappresenta un sistema di monitoraggio e controllo della tensione muscolare sviluppata nei muscoli degli arti inferiori. |
|||
|
|||
|
corrispondente all’area 3a di Brodmann; e da afferenze propriocettive provenienti dagli arti inferiori attraverso i cordoni laterali del midollo, il nucleo Z del tronco encefalico ed il talamo. |
|||
|
|



|
|